
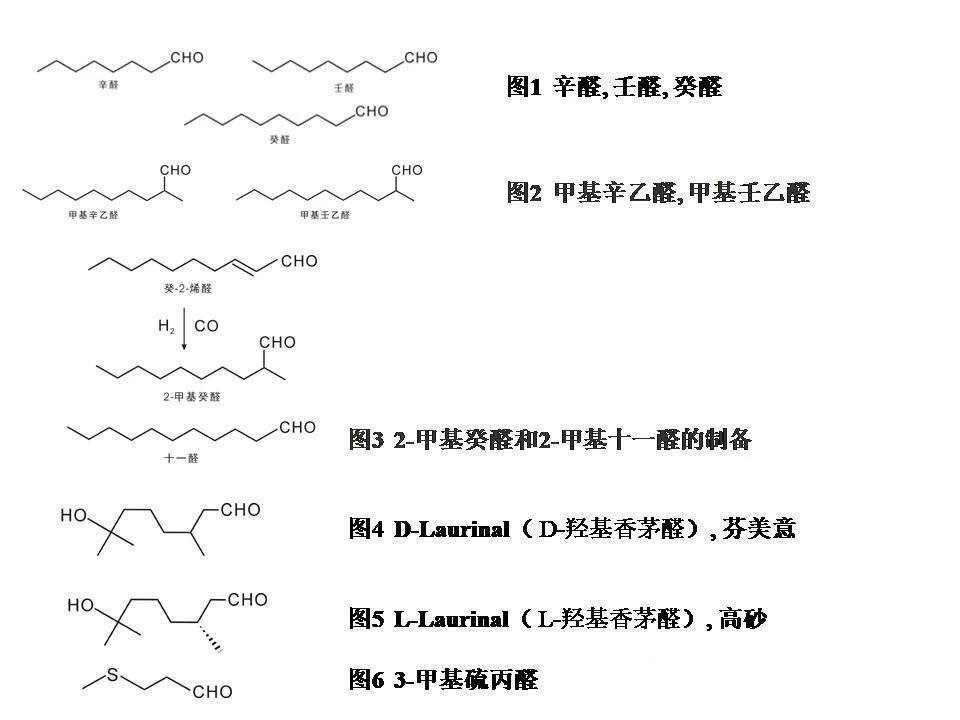

 玫瑰香精
玫瑰香精 清新花香香精
清新花香香精 姜花香精
姜花香精 葡萄柚香精
葡萄柚香精 花香香精
花香香精 洋甘菊香精
洋甘菊香精 小蒼蘭香精
小蒼蘭香精 茉莉香精
茉莉香精 水蜜桃香精
水蜜桃香精


服務(wù)熱線
177-5061-9273

香葉醇(英語:Geraniol,又稱牻牛兒醇)是一個非環(huán)單萜醇類化合物。它是玫瑰油、魯沙香茅油和香茅油等香精油的主要成分之一,也少量存在于天竺葵和檸檬中。常溫下為無色至黃色的油狀液體,具有溫和、甜的玫瑰花氣息,味有苦感。難溶于水,可溶于有機溶劑中。香葉醇及其酯廣泛用作日用香精和食用香精,是玫瑰系香精的主劑,用于配制日用產(chǎn)品和食品。
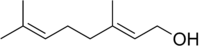
香葉醇的結(jié)構(gòu)式
工業(yè)制備
工業(yè)上用β-蒎烯高溫裂解生產(chǎn)月桂烯,然后使其在低溫及氯化亞銅和有機季銨鹽催化劑存在下與氯化氫加成,得到月桂烯的一級氯化物。將該氯化物與乙酸鈉和含氮堿(如三乙胺)共熱,得香葉醇、橙花醇和少量芳樟醇的乙酸酯混合物,經(jīng)皂化反應(yīng)水解,再蒸餾并仔細(xì)分餾便可得到香葉醇。
天然來源
可以提取出香葉醇的植物有:
唇形科:香薷 Elsholtzia ciliate (Thunb.) Hyland.,全草

香薷,中藥名。為唇形科、香薷屬植物,直立草本
百合科:大蒜 Allium sativum L.,揮發(fā)油
蕓香科:蕓香草 Cymbopoyon distans (Nees) A. Camus,揮發(fā)油

蕓香草
牻牛兒苗科:牻牛兒苗 Erodium stephanianum,揮發(fā)油

牻牛兒苗
樟科:月桂 Laurus nobilis L.,葉

月桂
蕓香科:九里香 Murraya paniculata (L. ) Jack.,葉

九里z香
薔薇科:玫瑰 Rosa rugosa Thunb.,揮發(fā)油
松藻科:刺松藻 Codium fragile (Sur.) Har.,全藻

刺松藻
香葉醇也是山梨酸代謝產(chǎn)物之一。
香葉醇作為基礎(chǔ)原料合成其他產(chǎn)物
香葉醇去掉羥基后衍生出的基團稱為香葉基(英文:Geranyl),也稱牻牛兒基。香葉基焦磷酸(即牻牛兒基焦磷酸,GPP)和香葉基香葉基焦磷酸(即牻牛兒基牻牛兒基焦磷酸,GGPP)是膽固醇生物合成過程中的重要中間產(chǎn)物。
香葉醇在酸性溶液中被轉(zhuǎn)化為環(huán)狀單萜α-松油醇。在銅催化劑存在下加熱,香葉醇可以重排成香茅醛,香茅醛經(jīng)還原可得到香茅醇。
香葉醇具有一定的毒性。大鼠口服半數(shù)致死量為3.6g/kg。
香葉醇在調(diào)香中的應(yīng)用
香葉醇及其酯類衍生物被廣泛用作日用香精,是玫瑰系香精的主劑,可用于草莓、桃、蘋果、橘子、菠蘿等果香型香精和生姜、肉桂等香型食用香精。也用于制取香葉醇酯(如乙酸香葉酯、甲酸香葉酯)、檸檬醛、香草醛、香草醇、維生素A、紫羅蘭酮和羥基香草醛,治療慢性支氣管炎,用作驅(qū)蚊劑成分、抗菌劑、香煙添加劑。
理化性質(zhì)
熔點-15°C
沸點229-230°C(lit.)
密度0.879g/mL at 20°C(lit.)
蒸氣密度5.31(vs air)
蒸氣壓~0.2mmHg(20°C)
FEMA2507|GERANIOL
折射率n20/D1.474(lit.)
閃點216°F
儲存條件2-8°C
溶解度water: soluble 0.1g/L at 25°C
形態(tài)Liquid
酸度系數(shù)(pKa)14.45±0.10(Predicted)
比重0.878~0.885(20/4℃)
顏色Clear colorless to pale yellow
水溶解性PRACTICALLY INSOLUBLE
JECFANumber1223
Merck14,4403
BRN1722456
穩(wěn)定性Stable. Combustible. Incompatible withstrong oxidizing agents.
InChIKeyGLZPCOQZEFWAFX-JXMROGBWSA-NCAS
數(shù)據(jù)庫106-24-1(CASDataBaseReference)
NIST化學(xué)物質(zhì)信息2,6-Octadien-1-ol,3,7-dimethyl-,(E)-(106-24-1)
EPA化學(xué)物質(zhì)信息trans-Geraniol(106-24-1)
香氣概述
香葉醇的氣味是盛開的玫瑰氣味,在香水工業(yè)中是主要的增甜劑,也是假玫瑰精油的主要成分。因此很多無良商家用天竺葵冒充玫瑰精油。香薰香葉醇就好像炙熱的愛情,激情火辣不失嬌媚,是勇往直前的愛情勇士。香葉醇的特性突出,抗微菌的能力是所有單萜醇里最好的,因此病毒感染的皮膚皰疹使用反式牻牛兒醇很有效。
來源
香葉醇主要來源于天竺葵,玫瑰,玫瑰草、爪哇香茅、蜂香薄荷,在高等級的佛手柑、快樂鼠尾草、芫荽、尤加利、永久花、薰衣草、檸檬草、香蜂草、百里香、野橘、依蘭依蘭精油中也含有少量的香葉醇。
含量分析
醇的總含量(以香葉醇計)按總醇量測定法(OT-5)測定。所取試樣量為1.2 g。計算中的當(dāng)量因子(e)取77.13。
毒性
ADI 0~5mg/kg(CE)。LD50 3600mg/kg(大鼠,經(jīng)口)。GRAS(FDA§182.60,2000)。
使用限量
FEMA(mg/kg):軟飲料2.1;冷飲3.3;糖果10;焙烤食品11;布丁類2.0;膠姆糖0.80~2.9;裱花層1.0。
化學(xué)性質(zhì)
無色至黃色油狀液體。具有溫和、甜的玫瑰花氣息,味有苦感。溶于有機溶劑,微溶于水。
用途
廣泛用于花香型日用香精,也可制成酯類香料,入藥用于抗菌和驅(qū)蟲
用途
GB2760—96規(guī)定為允許使用的食用香料。主要用以配制蘋果、桃、杏、李、草莓、檸檬、姜、肉桂、肉豆蔻和蜂蜜等香精。用途是一種天然香精香料,可用來制作香水,也可用作合成維生素E的原料
用途
是重要香料之一。常與香茅醇、苯乙醇共用,是各類玫瑰香精和配制香葉油的基本香料,廣泛應(yīng)用于花香香精,特別是玫瑰。也可限量用于晚香玉、香羅蘭、金合歡、Chemicalbook橙花、紫羅蘭、桂花、依蘭、香石竹、玉蘭等香精中。還可適量用于蘋果、杏子、草莓、懸鉤子、李子、桃子、蜂蜜、櫻桃等香型的食用香精中。
用途
香葉醇為玫瑰系香精的主劑,又是各種花香香精中不可缺少的調(diào)香原料,也可看作增甜劑,還可用于配制食品,香皂,日用化妝品香精。從天然精油中制取的產(chǎn)品,配制食品,日用化妝香精用時,含香葉醇大于90%,配制香皂香精用時,含香葉醇大于80%。工業(yè)品香葉醇和橙花醇是制造香草醇,香草醛、檸檬醛、羥基香草醛、紫羅蘭酮和維生素A的原料,用香葉醇合成的各種酯,也是很好的香料。
香葉醇的生物合成
1、工程大腸桿菌高效轉(zhuǎn)化香葉醇
單萜香葉醇(3,7-二甲基- 2,6 -辛二烯-1-醇的反式異構(gòu)體),從花中釋放出來,廣泛應(yīng)用于香水、制藥等行業(yè)。香葉醇作為汽油的替代品,其吸濕性低、能量含量高、揮發(fā)性較低,優(yōu)于乙醇。香葉醇是由香葉酰二磷酸(GPP)合成的,GPP在植物中由甲羥戊酸(MVA)途徑或甲基赤蘚醇磷酸途徑合成。然而,雖然香葉醇的經(jīng)濟價值很高,但如果它只能從植物中自然提取,濃度很低,將極大限制了其開發(fā)和應(yīng)用。因此,通過微生物宿主的代謝工程可以產(chǎn)生大量的增值產(chǎn)品。與植物不同,微生物通常不攜帶一種特定的GPP合酶(GPPS),也不能制造單萜,只有少數(shù)釀酒酵母菌能夠制造單萜(5 mg/L)。近年來,香葉醇已成功地在大腸桿菌和釀酒酵母中異源合成。金合歡基二磷酸合酶(FPPS)的突變允許GPP在含有單萜合酶的重組微生物中釋放用于單萜合成。最近的一項研究表明,在沒有外源香葉醇合成酶的情況下,突變的FPPS在酵母中積累GPP,可能通過內(nèi)源性去磷酸化作用使香葉醇形成。即使在大腸桿菌中沒有特定的GPPS或突變的FPPS,也可以通過過表達(dá)羅勒屬(Ocimum basilicum)香葉醇合成酶(GES)來生成香葉醇,盡管GPP的釋放機制尚不清楚。通過FPPS突變體與釀酒酵母GES的共同過表達(dá),培養(yǎng)7天后獲得5 mg/L香葉醇。通過過表達(dá)MVA途徑的關(guān)鍵限速酶,同時攜帶調(diào)控基因MAF1和GES的釀酒酵母在培養(yǎng)48 h后,香葉醇的產(chǎn)量進一步提高到36.04 mg/L。到目前為止,香葉醇脫氫酶突變體大腸桿菌在培養(yǎng)48 h后,通過全MVA途徑和GES產(chǎn)生的香葉醇最大值為182.5 mg/L。然而,對于工業(yè)應(yīng)用來說,滴度仍然太低。
此外,香葉醇通常會在芳香植物中轉(zhuǎn)化為其他萜類物質(zhì),從而影響蒸餾精油的品質(zhì)。香葉醇脫氫酶催化柔花香葉醇轉(zhuǎn)化為反式檸檬醛。類似地,一些葡萄酒酵母可以改變游離萜類成分,盡管它們只有有限的能力來生產(chǎn)單萜類化合物。釀酒酵母能夠在OYE2酶的催化下將香葉醇轉(zhuǎn)化為香茅醇,而ATF1醇乙酰轉(zhuǎn)移酶參與了釀酒酵母發(fā)酵過程中香葉醇的乙酰化。與酵母的廣泛研究不同,香葉醇在大腸桿菌中的生物轉(zhuǎn)化很少被提及,直到最近,香葉醇在大腸桿菌中被YjgB酶脫氫和異構(gòu)化成其他香葉醇(橙花醇、神經(jīng)醛和香葉醇)。
因此,Liu等創(chuàng)造了一種有效的香葉醇生物合成菌株,并開發(fā)了一種新的高效發(fā)酵策略來提高香葉醇的產(chǎn)量。此外,我們還觀察到在大腸桿菌中香葉醇和醋酸香葉醇之間的相互轉(zhuǎn)化。對乙酸香葉酯水解成香葉醇的機理進行了研究和控制,進一步提高香葉醇的產(chǎn)量。
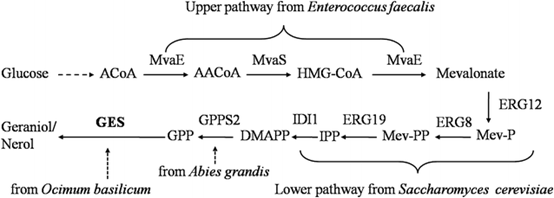
圖1 通過本研究中使用的甲羥戊酸(MVA途徑)產(chǎn)生香葉醇。參與該途徑的酶:MvaE、糞腸桿菌乙酰CoA: 乙酰基轉(zhuǎn)移酶/HMG-CoA還原酶;MvaS,糞腸桿菌HMG-CoA合成酶;ERG12,釀酒酵母甲羥戊酸激酶;ERG8,釀酒酵母磷酸丙二酸激酶;ERG19,釀酒酵母甲羥戊酸焦磷酸脫羧酶;IDI1,釀酒酵母IPP異構(gòu)酶;GPPS2,大葉香葉酰基二磷酸合成酶;GES, O. basilicum 合成酶被優(yōu)化為大腸桿菌的首選密碼子使用。途徑中間體:A-CoA,乙酰coa;AA-CoA acetoacetyl-CoA;β- hydroxymethylglutaryl-CoA;Mev-P,甲羥戊酸5-磷酸;Mev-PP,甲羥戊酸式焦磷酸鈉;IPP焦磷酸異戊烯基,DMAPP焦磷酸二甲基烯丙基,GPP香葉酰二磷酸
2、谷氨酸棒狀工程桿菌生產(chǎn)香葉醇
香葉醇是一種單萜醇,在食品、化妝品和保健中有多種應(yīng)用。谷氨酸棒狀桿菌是一個潛在的平臺萜類生產(chǎn),因為它承載甲基赤蘚糖醇磷酸途徑。為了將谷氨酰胺C. glutamicum轉(zhuǎn)化為香葉醇,分別表達(dá)了兩種不同的截斷香葉醇合成酶(GESs),表達(dá)該截斷香葉醇合成酶的菌株LX02 (tVoGES)產(chǎn)生0.3 mg/L的香葉醇。然后,將3個香葉酰二磷酸合成酶(GPPSs)與tVoGES聯(lián)合表達(dá),以提高香葉醇的產(chǎn)量。香葉醇的產(chǎn)量均高于菌株LX02。菌株LX03與tVoGES共表達(dá)ERG20F96W-N127W (ERG20WW)的產(chǎn)量最高,為5.4 mg/L。隨后,通過共同過表達(dá)1-脫氧-d -木酮糖-5-磷酸合酶(dxs)和異戊基二磷酸異構(gòu)酶(idi),菌株LX03的產(chǎn)量進一步提高到12.2 mg/L。最后通過發(fā)酵優(yōu)化,香葉醇的產(chǎn)量提高到15.2 mg/L。據(jù)我們所知,這是谷氨酰胺工程生產(chǎn)香葉醇的首次報道,可為其他單萜的生產(chǎn)研究提供參考。
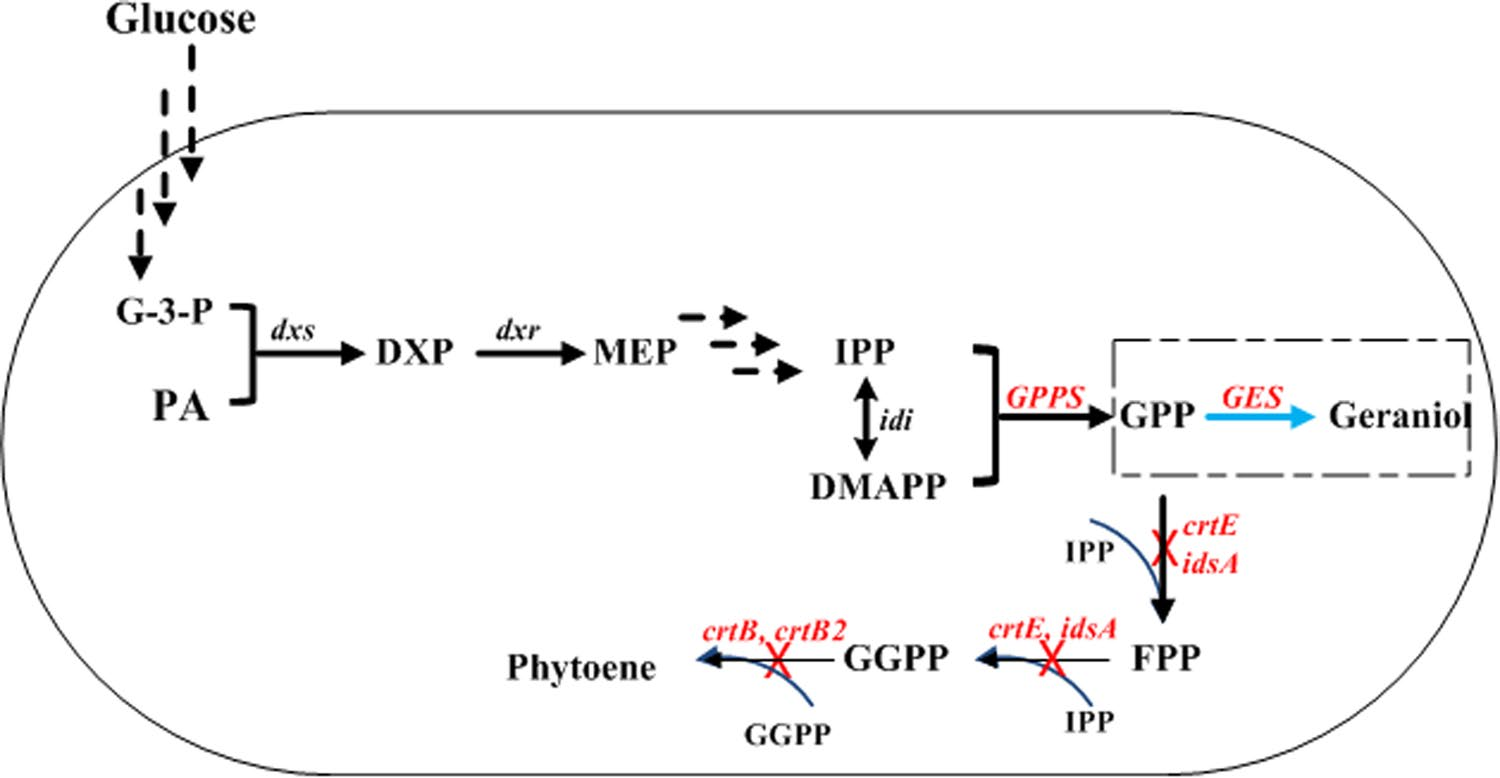
圖2 谷氨酰胺棒狀桿菌中香葉醇的生物合成途徑(黑色箭頭表示內(nèi)源性途徑,藍(lán)色箭頭表示外源性香葉醇合成步驟)。甘油醛3 -磷酸G-3-P;PA,丙酮酸;DXP 1-脫氧-d-木酮糖-5-磷酸;MEP,2-C-甲基-d-赤蘚醇-4-磷酸;IPP,異戊烯基焦磷酸;二甲基丙烯基焦磷酸 DMAPP;GPP,香葉基二磷酸;FPP,法尼基焦磷酸;香葉香葉基焦磷酸 GGPP;GES,編碼香葉醇合酶;GPPS,編碼香葉酰焦磷酸合酶;dx,編碼1-脫氧-d-木酮糖-5-磷酸合酶;Idi,編碼焦磷酸異戊烯基異構(gòu)酶;編碼植物烯合酶的crtB和crtB2;以及編碼GGPPS的idsA和crtE)
香葉醇的藥理活性
1、香葉醇的藥理作用綜述
香葉醇(Geraniol,簡稱GE)是一種非環(huán)類異戊二烯類單萜類化合物,從香樟、纈草等多種芳香植物的精油中分離得到。從植物分離中提取的香葉醇來源有限,不能滿足香精香料工業(yè)的巨大需求,需要通過生物技術(shù)最大限度地提高香葉醇的產(chǎn)量。香葉醇的多種活性表明,香葉醇可以作為一種很有前途的藥物來治療各種疾病。為了評價香葉醇在臨床試驗中的應(yīng)用潛力,本文對香葉醇的藥理作用進行了綜述。從PubMed、ScienceDirect、Springer和Wiley數(shù)據(jù)庫檢索的出版物被收集并總結(jié)了過去6年的文獻(xiàn)。然后,從香葉醇的抗腫瘤、抗炎、抗氧化、抗菌、保肝、保心、保神經(jīng)等藥理作用等方面探討了香葉醇的潛在應(yīng)用。因此,本文旨在為香葉醇的藥理活性提供證據(jù),為香葉醇作為一種臨床應(yīng)用候選藥物的進一步開發(fā)提供依據(jù)。
(1)、細(xì)胞毒性和抗腫瘤活性
GE對各種類型的癌癥具有細(xì)胞毒性和抗腫瘤活性。結(jié)腸癌是全球第四大癌癥致死原因。最近的一項研究表明,GE能顯著抑制結(jié)腸癌細(xì)胞株colo-205的細(xì)胞生長,IC50值為20μM。DAPI染色顯示GE可誘導(dǎo)細(xì)胞凋亡,Western blot檢測證實GE可上調(diào)Bax,下調(diào)Bcl-2。Bax是一種促凋亡的調(diào)節(jié)因子,Bcl-2是一種抗凋亡的調(diào)節(jié)因子。此外,GE誘導(dǎo)colo-205細(xì)胞的DNA損傷和細(xì)胞周期阻滯。子宮內(nèi)膜癌是世界范圍內(nèi)女性常見的癌癥。尋找治療子宮內(nèi)膜癌的新藥物是一項值得研究的課題。最近的研究表明,GE治療和預(yù)處理可通過抑制n-甲基-n'-硝基-n-亞硝基胍(150 mg/kg)誘導(dǎo)的雌性Wistar大鼠模型的癌基因和激活抑癌基因來抑制子宮內(nèi)膜癌。GE的抑制作用在預(yù)處理組更為有效。Real-time PCR結(jié)果顯示GE抑制K-ras、MAPK、PI3K和β-catenin的轉(zhuǎn)錄,Western blot結(jié)果顯示GE增加磷酸酶、PTEN、孕酮受體和E-cadherin蛋白的表達(dá)。MAPKs包括JNK、p38和ERKs。K-ras、MAPK、PI3K和β-catenin是癌基因,PTEN、孕酮受體和E-cadherin是腫瘤抑制基因。從血管瘤中分離得到的內(nèi)皮瘤細(xì)胞株eEND2容易生成血管。最近的一項研究顯示,GE對內(nèi)皮瘤細(xì)胞株eEND2具有抗血管生成作用。GE抑制內(nèi)皮樣eEND2細(xì)胞的遷移活動,同時抑制PCNA的表達(dá)。此外,GE可阻斷VEGF/VEGFR-2信號的轉(zhuǎn)導(dǎo)。VEGF促進癌細(xì)胞的血管生成。對血管生成的影響在體內(nèi)得到驗證:GE給BALB/c小鼠通過減弱VEGFR-2表達(dá),減少ki67陽性細(xì)胞和cd31陽性微血管的數(shù)量。
原發(fā)性肝癌是全球第三大癌癥死亡原因。從天然產(chǎn)物中分離得到的化合物對肝癌細(xì)胞具有細(xì)胞毒性作用。在Hep-G2肝細(xì)胞中,GE通過促進細(xì)胞凋亡發(fā)揮抑制作用。同時,GE抑制HMGCR蛋白和mRNA的表達(dá),導(dǎo)致膽固醇生成減少。在GE對肝細(xì)胞增殖的影響方面,GE可顯著抑制Hep-G2的增殖,其濃度可低至5μg/mL。此外,GE對二乙基亞硝胺相關(guān)肝癌的發(fā)生也有抑制作用,這可以通過降低血清中AST和ALT水平得到證實。此外,GE對PCNA和gst陽性面積比均有抑制作用。
肺癌是全世界男性的第一大死亡原因,女性的第三大死亡原因。裸鼠口服25、50和75 mmol/kg GE可通過增加細(xì)胞凋亡抑制肺癌細(xì)胞生長。后來的研究表明,GE下調(diào)HMGCR水平,降低膜結(jié)合Ras蛋白量,導(dǎo)致膽固醇生成減少。通過GE對甲羥戊酸途徑的抑制作用,A549腫瘤細(xì)胞的增殖受到抑制,細(xì)胞凋亡得到促進,而正常細(xì)胞中GE對細(xì)胞生存沒有任何抑制作用。
口腔癌占全世界癌癥的2-3%,但其存活率很低。因此,尋找一種治療口腔癌的新型候選藥物已迫在眉睫。在4NQO誘導(dǎo)的小鼠癌癥模型中,GE作為一種化學(xué)預(yù)防劑降低了口腔癌的發(fā)病率。在本模型中,GE通過NF-κB通路降低未成熟和成熟肥大細(xì)胞的密度,下調(diào)下游炎癥介質(zhì)(如TNF-α、IL-1β、COX-2和iNOS)的表達(dá)。另一項研究表明,在4NQO誘導(dǎo)的小鼠模型中,口服200 mg/kg GE的抑制作用有效,導(dǎo)致Nrf2通路和藥物代謝酶受到抑制。
在西方國家,胰腺癌是癌癥死亡的第四大原因。越來越多的證據(jù)表明,天然產(chǎn)物對胰腺癌細(xì)胞具有細(xì)胞毒性作用。在BXPC-3人胰腺癌細(xì)胞中,GE以劑量和時間依賴性的方式顯著抑制增殖和誘導(dǎo)凋亡。在吉西他濱(gemcitabine)治療前用GE預(yù)處理24小時,BXPC-3細(xì)胞表現(xiàn)出最大的抑制。本研究還發(fā)現(xiàn),GE通過誘導(dǎo)ROS的產(chǎn)生和下調(diào)酪氨酸激酶的磷酸化來抑制前列腺癌細(xì)胞的增殖。
前列腺癌仍然是美國人死亡的第一大原因。在前列腺腫瘤細(xì)胞中,GE通過下調(diào)轉(zhuǎn)錄因子E2F8的表達(dá)來抑制生長和生存。通過抑制這一關(guān)鍵的細(xì)胞周期控制因子,改變了參與細(xì)胞周期的基因表達(dá)譜,E2F8抑制通過誘導(dǎo)G2/M阻滯對細(xì)胞生長產(chǎn)生抑制作用。
在過去的幾十年里,皮膚癌的發(fā)病率一直在增加。因此,確定一種治療皮膚癌的新藥物是很有價值的。最近的報道表明,GE顯著抑制DMBA/TPA介導(dǎo)的瑞士白化病小鼠皮膚腫瘤的發(fā)生。GE抑制Ras/Raf/ERK1/2信號通路,誘導(dǎo)細(xì)胞凋亡。
(2)、抗炎和抗氧化活性
抑制炎癥反應(yīng)和氧化應(yīng)激可防止器官受損,尤其是大腦。GE可顯著降低TPA引起的LPO和炎癥反應(yīng)。進一步研究表明,GE促進炎癥細(xì)胞代謝,增加GSH含量,刺激抗氧化酶活性。此外,GE對p38MAPK活性的改變有抑制作用,減輕了TPA引起的NF-κB和COX-2的表達(dá)變化。另一項研究也表明GE可以通過抑制COX-2來抑制炎癥。GE通過刺激IL-10的產(chǎn)生,對帕米膦酸鈉誘導(dǎo)的炎癥反應(yīng)有抑制作用。此外,GE對單核細(xì)胞沒有毒性作用,也不影響TNF-α的產(chǎn)生。
一項研究對Wistar大鼠經(jīng)GE治療后的生化指標(biāo)和肝臟氧化應(yīng)激進行了測試,結(jié)果表明GE在芳香療法中可作為一種抗氧化劑。GE給藥與總膽固醇水平呈負(fù)相關(guān),但血糖、三酰基甘油蛋白和尿素水平無差異。GE處理的大鼠表現(xiàn)出GSH過氧化物酶和超氧化物歧化酶活性的增加,導(dǎo)致更多的過氧化物在肝臟中積累。GE可通過促進氧化應(yīng)激來上調(diào)ALT活性,但對AST活性無顯著影響。GE也被證明可以保護倉鼠免受ad引起的異常。100 mg/kg GE處理ad誘導(dǎo)的倉鼠,其機制包括減輕脂質(zhì)譜和內(nèi)皮功能標(biāo)志物的變化,促進抗氧化酶活性,減輕LPO標(biāo)志物和Nrf2表達(dá)的變化。組織學(xué)上,GE通過增加Nrf2活性負(fù)向影響氧化應(yīng)激。同時,GE具有抗炎、抗氧化作用。
潰瘍性結(jié)腸炎嚴(yán)重?fù)p害患者的生活質(zhì)量,迄今為止,沒有明確的標(biāo)準(zhǔn)護理被證明對這一適應(yīng)癥有效。GE能減少DSS引起的腸道損傷,這反映在結(jié)腸長度和大便一致性等疾病活動指數(shù)評分上。GE預(yù)處理對結(jié)腸組織中促炎細(xì)胞因子數(shù)量和髓過氧化物酶活性有抑制作用。GE下調(diào)下游目標(biāo)促炎酶的表達(dá),但上調(diào)抗氧化分子:抑制GSH和超氧化物歧化酶活性,阻止LPO和亞硝酸鹽的增加。
GE口服可通過減輕2-乙酰氨基芴誘導(dǎo)的氧化應(yīng)激、炎癥、增生和凋亡等組織損傷來保護肝癌組織。此外,GE能恢復(fù)受損的肝組織結(jié)構(gòu)。在分子機制上,GE降低了組織LPO,但提高了過氧化氫酶、谷胱甘肽過氧化物酶、谷胱甘肽還原酶和超氧化物歧化酶的活性。GE抑制了caspase-3、caspase-9、COX-2、NF-κB、PCNA、iNOS和VEGF的表達(dá),顯著降低了DNA的崩解。GE還可通過抑制內(nèi)臟脂肪、降低空腹血糖和血糖偏移來降低met誘導(dǎo)的炎癥和自由基損傷。
另一項研究表明,在DSS誘導(dǎo)的結(jié)腸炎小鼠模型中,GE可以預(yù)防結(jié)腸炎相關(guān)的生態(tài)干擾,并降低炎癥系統(tǒng),提示GE可以改善結(jié)腸炎的臨床癥狀。此外,研究表明,GE減弱了COX-2在腸壁的表達(dá)。采用脊髓損傷大鼠模型研究GE抗炎抗氧化作用的機制,結(jié)果表明,250 mg/kg/d GE處理大鼠4周后,可顯著降低脊髓含水量,提高Beattie和Bresnahan評分。此外,GE對炎癥和氧化應(yīng)激的抑制作用是通過減弱NF-κB和p38 MAPK通路介導(dǎo)的。
(3)、抑菌活性
白念珠菌是一種致人免疫功能低下的真菌致病菌,具有較高的致死率。最近的一項研究表明,GE治療可減弱白色念珠菌和非白色念珠菌的活性。口服GE對病原菌生物膜形成和菌絲形態(tài)形成有抑制作用。GE通過下調(diào)質(zhì)膜ATP酶活性和降低麥角甾醇水平來破壞細(xì)胞壁功能。GE通過促進鈣調(diào)神經(jīng)磷酸酶信號突變體的敏感性,同時增強對鈣調(diào)神經(jīng)磷酸酶過表達(dá)菌株的耐藥性來抑制真菌的增殖。此外,GE破壞線粒體功能,破壞鐵穩(wěn)態(tài),減輕遺傳毒性。另一項研究表明,GE對白色念珠菌的最低抑菌濃度為16μg/mL,且其抑菌作用呈濃度依賴性。形態(tài)干擾試驗表明,GE對偽菌絲和衣原體孢子的形成有抑制作用。紅色毛癬菌是慢性皮膚病中最常見的真菌之一。GE可通過其抗真菌活性治療皮膚的紅色結(jié)核菌感染。高濃度GE能抑制菌絲和真菌的生長,也能抑制分生孢子的萌發(fā)。GE主要通過胞內(nèi)物質(zhì)的泄漏和抑制麥角甾醇的生物合成來破壞胞壁和細(xì)胞膜。
(4)、護肝效應(yīng)
通過口服200 mg/kg GE可減輕蛋氨酸膽堿缺乏飲食誘導(dǎo)的大鼠非酒精性脂肪性肝炎。在該模型中,GE對肝臟的組織學(xué)評分、纖維化和凋亡均有抑制作用。在分子水平上,GE抑制血清中丙氨酸轉(zhuǎn)氨酶和天冬氨酸轉(zhuǎn)氨酶的活性。GE通過抑制肝臟線粒體ROS、線粒體電子傳遞鏈酶活性和線粒體DNA含量來保護肝臟線粒體功能。此外,GE能減弱丙二醛和3-硝基酪氨酸的形成,抑制肝臟中iNOS和細(xì)胞色素P450 2E1的表達(dá)。GE對肝臟炎癥有抑制作用,抑制TNF-α、IL-6的表達(dá)和髓過氧化物酶活性。蘇木精、伊紅染色和PCR結(jié)果顯示,在肺動脈高壓(PH)為70%的大鼠中,GE表現(xiàn)出肝保護作用,腹腔注射GE可通過提高TNF-α和IL-6的表達(dá)水平促進肝組織再生。同時,GE顯著抑制血清ALT水平,調(diào)節(jié)肝細(xì)胞功能恢復(fù)。GE是一種天然化合物,可以顯著促進肝臟再生,在PH模型大鼠中注射GE 100 mg/kg。在機制上,GE增加NF-κB、TNF-α、IL-6 mRNA和蛋白表達(dá)水平,抑制HSP27、HSP60蛋白表達(dá)水平。通過腹腔注射50 mg/kg GE可部分恢復(fù)h2o2誘導(dǎo)的氧化應(yīng)激模型大鼠的氧化性肝損傷。其機制是通過GE調(diào)節(jié)H2O2引起的丙二醛和谷胱甘肽的變化,并抑制肝臟過氧化氫酶活性。
(5)、護心效應(yīng)
GE可減輕AD模型小鼠的高脂血癥,降低冠心病的發(fā)生率。GE通過下調(diào)HMGCR和抑制脂肪生成來降低血脂。GE抑制LCAT,降低膽固醇酯水平。同時,GE處理可提高C反應(yīng)蛋白的活性。GE保護心血管系統(tǒng)免受高血脂對身體的傷害。特別是,GE治療降低血漿中總膽固醇和總甘油三酯的產(chǎn)生,并抑制肝臟脂肪酸、總脂質(zhì)和非皂化脂質(zhì)的生物合成。GE通過增加LDL mRNA和VLDL受體mRNA增加血清LDL攝取。最后,GE抑制甘油三酯的合成,降低脂肪酸合成。多年來,GE作為治療高脂肪飲食引起的內(nèi)皮細(xì)胞功能障礙的候選藥物,引起了人們的關(guān)注。此外,GE可下調(diào)血清硫代巴比妥酸反應(yīng)性物質(zhì)和主動脈ROS的產(chǎn)生。GE可通過抑制主動脈中NOX-2的表達(dá),保護高脂飲食誘導(dǎo)的血管內(nèi)皮功能障礙。GE抑制ROS水平,減少心肌缺血-再灌注過程中的損傷,促進心肌細(xì)胞功能和活力。在新生大鼠心室心肌細(xì)胞中,GE可降低內(nèi)源性ROS的產(chǎn)生,并對pAMPK水平產(chǎn)生正向調(diào)節(jié),同時降低pERK1/2水平(pAMPK和pERK1/2是調(diào)節(jié)細(xì)胞存活的關(guān)鍵因素)。可見,GE對心肌細(xì)胞具有保護作用。GE通過抑制氧化應(yīng)激和提高氧化應(yīng)激標(biāo)志物8-異前列腺素對糖尿病致心功能障礙具有心臟保護作用。
(6)、神經(jīng)保護效應(yīng)
在ACR誘導(dǎo)的果蠅神經(jīng)損傷模型中,GE表現(xiàn)出神經(jīng)保護作用,ACR引起的死亡率明顯降低。GE補充可減輕acr誘導(dǎo)的氧化應(yīng)激、線粒體功能障礙和神經(jīng)毒性。其機制是隨著解毒酶活性的增加而恢復(fù)谷胱甘肽和總硫醇水平。此外,GE與姜黃素同時給藥對膽堿能功能有負(fù)作用,乙酰膽堿酯酶活性降低。帕金森病是指中樞神經(jīng)系統(tǒng)長期退行性疾病,影響運動系統(tǒng)。在MPTP誘導(dǎo)的帕金森病(PD)小鼠模型中,GE預(yù)處理對PD引起的神經(jīng)系統(tǒng)損傷具有保護作用。GE口服可通過增加酪氨酸羥化酶免疫反應(yīng)性表達(dá)和降低α-突觸核蛋白(synuclein)表達(dá)來改善神經(jīng)肌肉功能障礙。GE可減輕mptp誘導(dǎo)的PD小鼠模型的運動行為和神經(jīng)營養(yǎng)因子不足。另一項研究表明,在MPTP誘導(dǎo)的PD小鼠模型中,給藥GE可調(diào)節(jié)Bcl-2/Bax比值,抑制細(xì)胞色素c和caspase-9的表達(dá),從而減輕神經(jīng)變性和運動紊亂。細(xì)胞色素c與心磷脂結(jié)合,釋放出線粒體并引發(fā)細(xì)胞凋亡。Caspase-9是一種啟動凋亡通路的酶。GE對轉(zhuǎn)基因PD模型果蠅PD臨床癥狀有抑制作用。GE通過降低GSH含量、增加蛋白質(zhì)羰基含量、LPO和GST活性來調(diào)節(jié)氧化應(yīng)激。GE顯著抑制α-突觸核蛋白表達(dá),導(dǎo)致果蠅PD臨床癥狀減弱。在脊髓損傷大鼠中,GE可通過給藥減輕神經(jīng)痛和功能損害。GE對NeuN陽性細(xì)胞有正向作用,但抑制損傷區(qū)膠質(zhì)纖維酸性蛋白的表達(dá),降低半胱天冬酶(caspase-3)的活性。此外,本研究還發(fā)現(xiàn),GE處理可降低急性脊髓損傷(SCI)大鼠丙二醛和3-硝基酪氨酸水平,促進核因子紅系2相關(guān)因子2和血紅素加氧酶1蛋白表達(dá),降低iNOS表達(dá)。綜上所述,GE對脊髓損傷恢復(fù)有積極作用,可減輕神經(jīng)性疼痛。因此,GE是治療脊髓損傷的候選藥物。
(7)、抗糖尿病效應(yīng)
糖尿病是一種代謝紊亂,表現(xiàn)為高血糖水平,并可能導(dǎo)致各種并發(fā)癥。在鏈脲佐菌素(STZ)誘導(dǎo)的糖尿病大鼠模型中,GE可用于治療糖尿病,GE給藥45 d可降低血糖和血紅蛋白HbA1C,并可恢復(fù)胰島素反應(yīng)。許多參與葡萄糖生產(chǎn)和利用的酶的活性被恢復(fù),導(dǎo)致碳水化合物代謝的改善和葡萄糖穩(wěn)態(tài)的恢復(fù)。在鏈脲佐菌素誘導(dǎo)糖尿病大鼠模型的另一項研究顯示,GE治療對糖尿病神經(jīng)病變有保護作用,提高了感覺和運動功能減少酶的活動(例如,復(fù)合物I - III,琥珀酸脫氫酶,和檸檬酸合成酶)在坐骨神經(jīng)細(xì)胞溶質(zhì)和選定的大腦區(qū)域。GE能降低胞質(zhì)鈣水平和乙酰膽堿酯酶活性。同時,GE可以抑制羰基和亞硝酸鹽蛋白。GE可通過抑制氧化應(yīng)激和降低高血糖來減輕糖尿病引起的另一并發(fā)癥,即糖尿病引起的心功能障礙。GE治療可提高心臟收縮功能,恢復(fù)高血糖所致心臟舒張功能。此外,GE通過阻止8-異前列腺素的增加來顯著抑制氧化應(yīng)激。GE可減輕糖尿病或多發(fā)性硬化(MS)小鼠的血管反應(yīng)性損傷,即GE以劑量依賴的方式改善糖尿病或MS暴露于甲乙二醛引起的血管過度收縮。該研究進一步證明,GE預(yù)孵育可改善血管收縮。此外,GE對電壓依賴和受體介導(dǎo)的鈣通道引起的血管收縮也有抑制作用。GE通過阻斷這些鈣通道減少受損血管的反應(yīng)。
(8)、其他效應(yīng)
在哺乳動物模型中,GE對哇吧因(ouabain)誘發(fā)的心律失常有抗心律失常作用。GE通過下調(diào)心臟收縮力來減輕哇吧因引起的心律失常。GE阻斷了瞬時外向鉀(K+)電流(59.7%)、非失活鉀(39.2%)和內(nèi)向整流鉀(33.7%),使動作電位持續(xù)時間增加了50%。GE還能降低左室壓(83%)和心率(16.5%)。GE可使哇吧因誘發(fā)的心律失常起病時間延長128%,靜息張力降低30%。
GE對胃和十二指腸粘膜有良好的抗?jié)兓钚浴E灌胃后,7.50 mg/kg GE可誘導(dǎo)GSH含量增加,髓過氧化物酶水平降低。此外,GE可使缺血/再灌注誘導(dǎo)的十二指腸潰瘍損傷減少71%,半胱胺誘導(dǎo)的十二指腸潰瘍減少68%。GE通過增強內(nèi)源性前列腺素和一氧化氮途徑等保護因子的活性來保護胃和十二指腸粘膜。這種保護機制還包括增加具有抗氧化特性的粘液分泌。GE增加降鈣素基因相關(guān)肽的釋放。GE對三硝基苯磺酸(TNBS)誘導(dǎo)的結(jié)腸炎損傷有保護作用。具體而言,GE治療TNBS誘導(dǎo)的大鼠結(jié)腸炎11天,臨床癥狀(如體重減輕、結(jié)腸水腫、潰瘍面積和結(jié)腸/脾臟質(zhì)量指數(shù))有所改善。GE和/或柳氮磺胺吡啶可抑制大鼠結(jié)腸炎模型中一氧化氮和脂質(zhì)過氧化物水平升高,恢復(fù)大鼠抗氧化活性。在分子水平上,GE通過降低半胱胺天冬酶caspase-3、細(xì)胞間黏附分子-1和髓過氧化物酶的水平,抑制細(xì)胞凋亡和炎癥細(xì)胞浸潤。細(xì)胞間粘附分子-1是一種跨膜蛋白,能夠穩(wěn)定細(xì)胞間的相互作用,促進白細(xì)胞內(nèi)皮細(xì)胞的轉(zhuǎn)運。髓過氧化物酶氧化降低細(xì)胞凋亡和炎癥的抑制。相反,GE的抗炎作用則與下調(diào)結(jié)腸前列腺素E2和IL-1β含量有關(guān)。
GE口服3周后對小鼠CUMS模型表現(xiàn)出類似抗抑郁作用,并可顯著縮短小鼠懸尾靜止時間。更特別的是,GE降低慢性不可預(yù)測輕度應(yīng)激(CUMS)誘導(dǎo)的促炎因子IL-1β水平,從而改善抑郁行為。GE處理對NF-κB通路的激活有抑制作用。此外,GE調(diào)節(jié)了核苷酸結(jié)合和包含3個炎癥小體的寡聚結(jié)構(gòu)域樣受體家族pyrin結(jié)構(gòu)域的表達(dá)。GE治療(100mg /kg b.w)可通過抑制轉(zhuǎn)錄因子NF-κB的表達(dá),從而阻礙組織重塑,從而減輕動脈粥樣硬化誘導(dǎo)的組織纖維化。NF-κB的弱表達(dá)可減輕氧化應(yīng)激和炎癥反應(yīng),對組織纖維具有保護作用。
GE可以通過殺死陰道毛滴蟲來治療陰道毛滴蟲。GE破壞核膜和細(xì)胞核,導(dǎo)致染色質(zhì)積累,細(xì)胞質(zhì)中出現(xiàn)大量空泡。核糖體減少,細(xì)胞器解體,細(xì)胞膜受損。有趣的是,GE并沒有表現(xiàn)出時間依賴性的抑制作用。
GE給藥100或200 mg/kg可減輕卵清蛋白致敏模型大鼠的過敏性哮喘。研究表明,GE對支氣管肺泡灌洗液中人嗜酸細(xì)胞激活趨化因子(eotaxin)水平及嗜酸性粒細(xì)胞浸潤有抑制作用。即GE干預(yù)降低了支氣管肺泡灌洗液中輔助型T細(xì)胞2(TH2)細(xì)胞因子,但刺激了輔助型T細(xì)胞1(TH1)細(xì)胞因子干擾素。此外,GE促進了核轉(zhuǎn)錄相關(guān)因子(Nrf2)蛋白的表達(dá),并強化了Nrf2定向的抗氧化途徑(如谷氨酸-半胱氨酸連接酶、超氧化物歧化酶和谷胱甘肽-S 轉(zhuǎn)移酶)。GE還增加了GSH的產(chǎn)量。在實驗小鼠模型中,GE在炎癥引起的疼痛后表現(xiàn)出抗傷害性感受活性。抗傷害性的活動是由于周圍神經(jīng)興奮性的降低。行為試驗表明,GE處理可減少醋酸引起的扭體次數(shù)。此外,GE可通過調(diào)節(jié)谷氨酸神經(jīng)傳遞減少小鼠舔爪時間。GE治療可通過抑制神經(jīng)遞質(zhì)和炎癥產(chǎn)生介質(zhì)來減輕疼痛。GE對急性乙醇誘導(dǎo)的潰瘍和慢性乙酸誘導(dǎo)的潰瘍也有胃保護和胃愈合作用。
最近,從天然產(chǎn)品中提取的純化合物被認(rèn)為是有潛力的補充和替代藥物,用于治療各種疾病。一般來說,單一化合物可以靶向DNA、mRNA、蛋白質(zhì),甚至是microRNA。近幾十年來,越來越多的證據(jù)表明,GE是一種純植物化合物,無不良反應(yīng),主要通過調(diào)節(jié)蛋白表達(dá)發(fā)揮多種活性,有望成為治療多種疾病的新型候選藥物。GE對癌細(xì)胞的細(xì)胞毒性作用表明,GE可以治療癌癥,降低癌癥患者的死亡率。GE的抗炎和氧化作用表明GE具有保護器官損傷和治療潰瘍性結(jié)腸炎的作用。GE的抗真菌活性表明其對真菌感染有一定的保護作用。GE的抗糖尿病作用說明GE具有治療糖尿病的作用。GE的抗傷害感受活性表明GE可以作為一種鎮(zhèn)痛藥用于臨床試驗。然而,GE的抗癌作用還需要在更多的動物模型中進行體內(nèi)研究,并在隨后的人類患者中進行研究,以證實GE對惡性腫瘤的抑制作用。此外,值得探討其藥理作用的分子基礎(chǔ)(如抗菌和抗心律失常活性)。缺乏有關(guān)GE的明確靶標(biāo)的信息將是反對GE在臨床試驗中應(yīng)用的一個原因。
2、香葉醇的抗腫瘤作用:調(diào)節(jié)腫瘤信號通路(綜述)
香葉醇是一種膳食單萜醇,存在于芳香植物的精油中。迄今為止,實驗證據(jù)支持香葉醇對乳腺癌、肺癌、結(jié)腸癌、前列腺癌、胰腺癌和肝癌等不同類型的癌癥有治療或預(yù)防作用,并揭示了其藥理作用的機制基礎(chǔ)。此外,香葉醇使腫瘤細(xì)胞對常用的化療藥物敏感。香葉醇控制多種代表腫瘤特征的信號分子和通路;香葉醇的這些作用抑制了腫瘤細(xì)胞獲得抗腫瘤藥物適應(yīng)性耐藥性的能力。在這篇綜述中,我們強調(diào)香葉醇是一種很有前途的化合物或化學(xué)基團,可用于開發(fā)一種安全有效的多靶點抗癌藥物。我們總結(jié)了香葉醇對癌細(xì)胞靶分子和通路的影響。本文綜述了香葉醇研究面臨的挑戰(zhàn)和前景,以及香葉醇在未來臨床研究中的應(yīng)用。
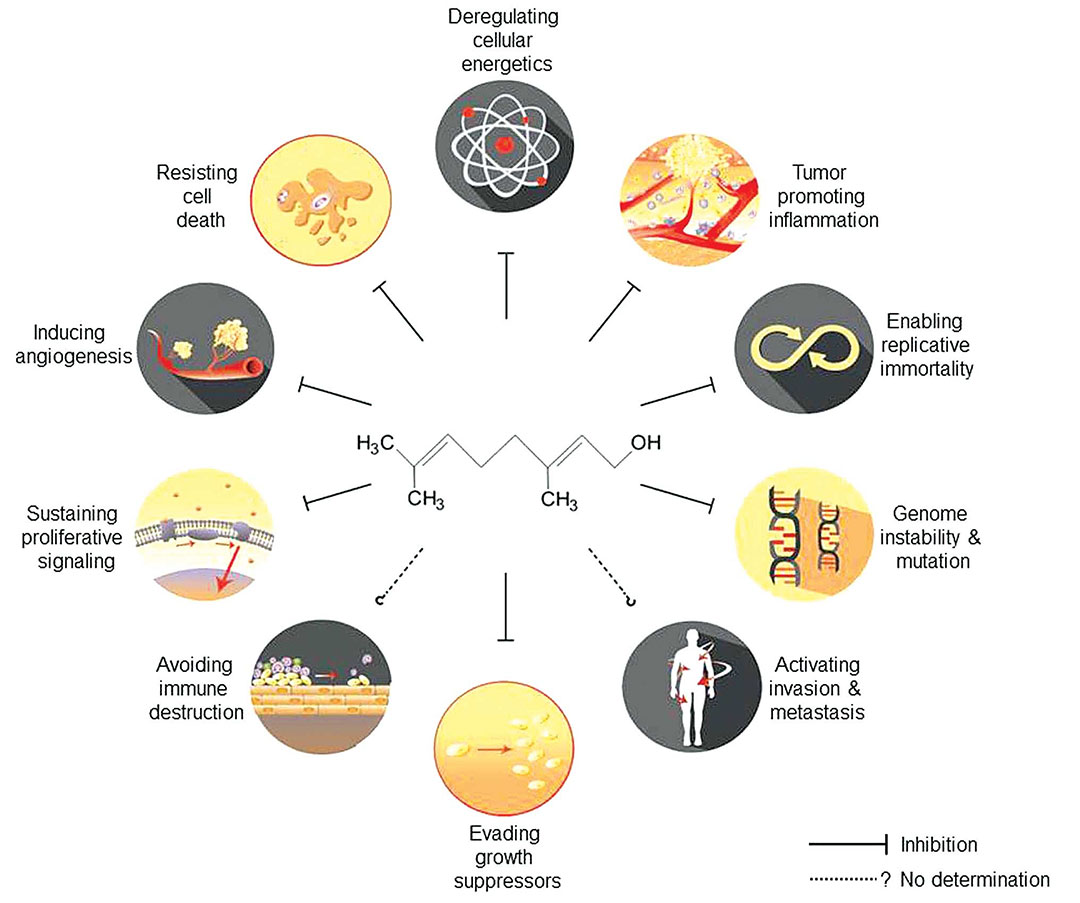
圖3 香葉醇對癌癥的多重作用
香葉醇已被發(fā)現(xiàn)對多種癌癥有效,包括乳腺癌、肺癌、結(jié)腸癌、前列腺癌、胰腺癌、皮膚癌、肝癌、腎癌和口腔癌(圖2)。本文總結(jié)了香葉醇對各種癌癥的治療作用的分子機制。在癌癥的特征中,使復(fù)制的永生和抵抗細(xì)胞死亡似乎是香葉醇最主要的目標(biāo)。然而,需要強調(diào)的是,這可能是研究偏好或發(fā)表偏好:例如,香葉醇是否影響避免免疫破壞或激活入侵和轉(zhuǎn)移尚未報道。給出了香葉醇在體外(細(xì)胞培養(yǎng))和體內(nèi)(異種移植或化學(xué)致癌模型)的有效劑量范圍。
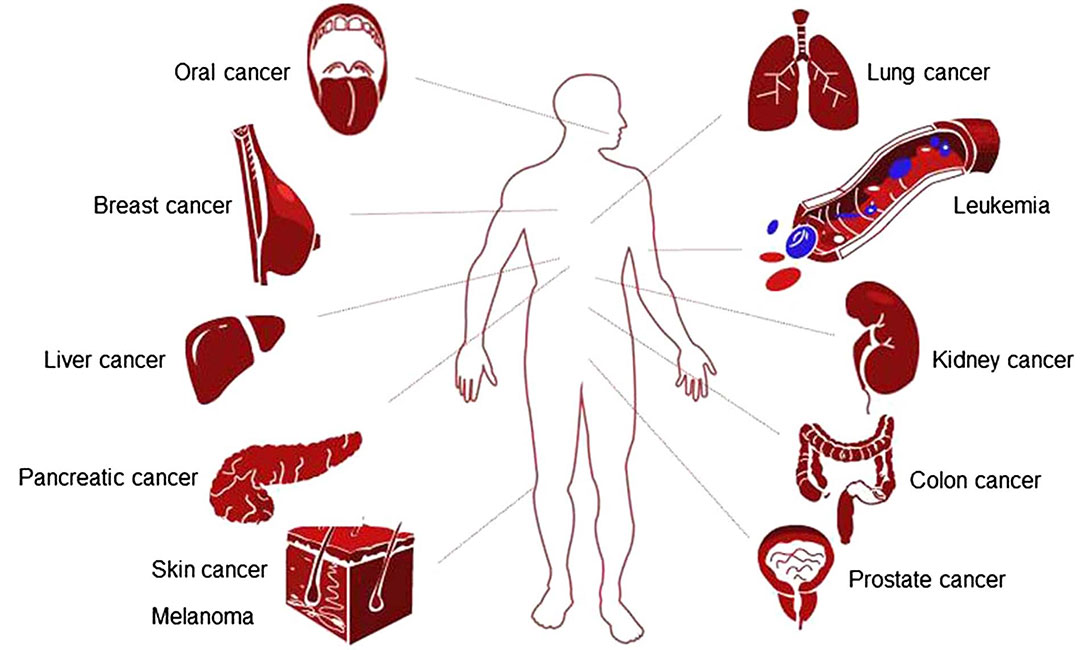
圖4 對香葉醇敏感的癌癥類型
參考文獻(xiàn)
[1] Liu, W., Xu, X., Zhang, R. et al. Engineering Escherichia coli for high-yield geraniol production with biotransformation of geranyl acetate to geraniol under fed-batch culture. Biotechnol Biofuels 9, 58 (2016). https://doi.org/10.1186/s13068-016-0466-5
[2] Li, M., Xu, S. & Lu, W. Engineering Corynebacterium glutamicum for Geraniol Production. Trans. Tianjin Univ. 27, 377–384 (2021). https://doi.org/10.1007/s12209-020-00252-w
[3] Yu Lei, Peng Fu, Xie Jun, Peng Cheng. PharmacologicalProperties of Geraniol – A Review. Planta Med 2019; 85(01): 48-55.DOI:10.1055/a-0750-6907.
[4] Cho, M., So, I., Chun, J.N., & Jeon, J. (2016). The antitumor effects of geraniol: Modulation of cancer hallmark pathways (Review). International Journal of Oncology, 48, 1772-1782. https://doi.org/10.3892/ijo.2016.3427.